Anterior dentition of Indohyus indirae
3D models of the skull of Miocaperea
3D models of placental mammal turbinal complex
3D GM dataset of bird skeletal variation
Skeletal embryonic development in the catshark
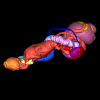
Bony connexions of the petrosal bone of extant hippos
bony labyrinth (11) , inner ear (10) , Eocene (8) , South America (8) , Paleobiogeography (7) , skull (7) , phylogeny (6)
Lionel Hautier (22) , Maëva Judith Orliac (21) , Laurent Marivaux (15) , Pierre-Olivier Antoine (12) , Bastien Mennecart (12) , Renaud Lebrun (10) , Rodolphe Tabuce (10)